yogabook / movement physiology / muscle
Contents
- 1 The muscle
- 1.1 General information and structure
- 1.1.1 Smooth and striated muscles
- 1.1.2 Muscle fibres
- 1.1.3 Sarcomere
- 1.1.4 Equilibrium length
- 1.1.5 Cross-bridge cycle
- 1.1.6 Titin
- 1.1.7 Sarcomere length
- 1.1.8 bilateral strength deficit
- 1.1.9 general strength deficit
- 1.1.10 Forms of contraction
- 1.1.11 Heads and bellies
- 1.1.12 The type of fibre
- 1.1.13 Feathering
- 1.2 Contraction
- 1.3 Articulation
- 1.4 Training
- 1.4.1 Incentives for muscle building
- 1.4.2 Trainingsparameter
- 1.4.3 Supercompensation theory
- 1.4.4 Combination exercises
- 1.4.5 Endurance sports versus weight training and muscle building
- 1.4.6 Further information
- 1.4.7 Intensity levels of the training
- 1.4.8 training zones
- 1.4.9 Fat and carbohydrates
- 1.4.10 Muskelkater (Munich Consensus Statement 1b)
- 1.4.11 Muscle strength
- 1.4.12 Muscle performance
- 1.4.13 Muscle failure
- 1.4.14 Eccentric muscle failure
- 1.4.15 Isometric muscle failure
- 1.4.16 concentric muscle failure
- 1.4.17 longitudinal muscle adaptation
- 1.4.18 Pre-activation
- 1.5 Sarcopenia
- 1.6 Dynapenia
- 1.7 Myogelosis
- 1.8 Arthrogenic muscle inhibition (AMI)
- 1.9 Stretching
- 1.10 Slackening points and insufficiencies
- 1.11 Torque
- 1.1 General information and structure
The muscle
General information and structure
The muscle (from the Latin musculus: little mouse) is an organ capable of contraction and relaxation, which attaches to bones or soft tissue structures such as connective tissue to enable movements of the body, both in the movement apparatus and in other organs, such as the ligaments.
- in the peristalsis of digestion (wave-like contractions for the transport of chyme or faeces)
- as sphincter (sphincter muscle)
- as pump (heart)
- as a regulator of vascular tension, e.g. in blood vessels.
Smooth and striated muscles
While the passive movement apparatus accounts for around 10% of body weight, the muscles make up around 40%, in athletes up to 65%, in newborns 20%. A distinction is made between:
Smooth muscles:
is found in the walls of all cavity organs except the heart. It lacks the microscopically visible transverse striation of the skeletal musculature and cannot be contracted arbitrarily. It controls itself or is subject to the influence of hormones, neurotransmitters, other messenger substances, the vegetative or enteric nervous system.
striated muscles
These include heart muscles and skeletal muscles, muscles of the tongue, larynx and diaphragm. Of all these, only the heart is not subject to voluntary control. In the skeletal musculature, the insertion points are usually referred to as origin and approach, although the origin is the point/area closer to the centre of the body, at least in the limbs. This often coincides with the view that the origin is the punctum fixum (fixed point) and the insertion is the punctum mobile (moving point), for which, however, the rectus femoris in sit-ups is a counter-example. The origin is also not clear, as shown by the biceps brachii or triceps brachii. In the shoulder girdle, the attachment to the trunk is usually referred to as the origin when the muscle moves the scapula (trunkoscapular musculature) or the arm (trunkohumeral musculature), and that at the scapula is referred to as the origin if the muscle moves the arm from there (scapulohumeral musculature). If a muscle attaches to a bone end (epiphysis), the transition is slightly cartilaginous, which provides minimal elasticity; if it attaches to a bone shaft (diaphysis), minimal damping is provided by the interweaving of the collagen fibres of the tendon with the elastic fibres of the connective tissue outer layer (stratum fibrosum) of the periosteum (bone skin).
Muscle fibres
A skeletal muscle consists of several (secondary) bundles of muscle fibres separated by a fascia (perimysium). The secondary bundles consist of several primary bundles. The secondary bundles are surrounded by perimysium externum, which extends inwards with septa and surrounds the primary bundles as perimysium internum. The individual approx. 20 µm wide muscle fibres (myocytes), which make up a primary bundle approx. 40 – 80 µm wide (similar to a human hair), are each surrounded by endomysium.
The muscle fibres are usually up to 15 cm long, in the case of M. sartorius also 30-40 cm long cells: myocytes. The myocytes have several cell nuclei that dominate a certain „area of influence“ (MND: myonuclear domain). The MND can be increased through strength training. Myocytes contain thousands of thread-like, parallel myofibrils as cell organelles. The myofibrils consist of many microfilaments arranged one behind the other, the actual contractile elements of the muscle. When stained under the microscope, these look like they are lying side by side along the length of the myofibril, hence the name striated muscle.
The maximum force of skeletal muscles is around 5 kg or 50 N per square centimetre on average, which is around 500N for biceps. Their maximum shortening compared to the longest state in vivo is around 40%. In vitro, the muscle is slightly longer at rest than in vivo.
Sarcomere
The smallest contractile functional unit of a muscle is the sarcomere. In most muscles, there are more than 1000 sarcomeres in a row in the myofibrils. Depending on the degree of contraction of the muscle (and thus depending on the innervation), the sarcomeres are between a minimum of just over 1 µm and a maximum of just under 5 µm long. Its average resting length is 2-2.5 µm. In vivo, a sarcomere barely reaches more than 4 µm in length. They are bordered longitudinally by Z-discs on which actin filaments are suspended, which have a larger number of possible binding sites for another protein, myosin, in the longitudinal direction. The myosin filaments are attached to another disc, the M-disc, in the (longitudinal) centre of the sarcomere, so that the myosin filaments protrude into the spaces between the actin filaments. The myosin filaments each consist of approx. 300 myosin proteins with tails twisted together in pairs, each with a neck and a head. This is why they are also referred to as thick filaments, while the much thinner actin filaments are called thin filaments. The actin filaments are in the form of a double helix, to which an attached tropomyosin thread is attached at rest in such a way that it covers the binding sites for myosin. The troponin attached to the tropomyosin can swivel the filament to the side so that the binding sites are exposed. At rest, the myosin head is at a low-energy angle of 45° to the tail (and to the actin). An ATP molecule is attached to it. For a more detailed description of the sarcomere, see Wikipedia.
Equilibrium length
A muscle has a shorter length in vitro than in the context of the movement apparatus. The length that occurs without innervation is referred to as the equilibrium length. If the muscle is stretched beyond this length, passive restoring forces occur that increase exponentially in the direction of greater sarcomere length and exceed the maximum achievable isometric muscle force. This restoring force is based on the elastic modulus of the titin, which connects the myosin with the Z-disc of the sarcomere.
Cross-bridge cycle
At rest, the myosin head is at a low-energy angle of 45° to the tail (and to the actin). An ATP molecule is attached to it. By hydrolysis of the ATP to ADP plus a phosphate residue, the head is positioned at an angle of 90° to the tail. Mg is involved in this reaction. If Ca now flows in after a nerve impulse from the sarcoplasmic reticulum and binds to troponin, this swings the tropomyosin thread to the side so that the myosin can bind to the nearest binding site of the actin. This binding is called a cross-bridge. Now the myosin head releases the still bound ADP and the phosphate residue, causing the neck to abruptly change its angle back to the low-energy 45° position. This movement is known as a power stroke. The empty binding site for ATP on the myosin head is reoccupied with ATP, whereupon the head detaches from the actin and returns to the 90° position, from which it can bind again if a binding site released by the tropomyosin is nearby. At the same time, the Ca is released from the troponin, whereupon the tropomyosin blocks the binding sites again. The resulting cycle is called a cross-bridge cycle. Cross-bridge cycles last between about 10 and 100 ms. Around 50 cycles can be completed within one second, which can shorten the muscle by half if it was previously at a long sarcomere length. How long cross-bridge cycles are carried out in succession depends on the Ca influx and thus on the innervation. The membrane of the sarcoplasmic reticulum contains channels for the sudden release of Ca for the initiation of cross-bridge cycles as well as Ca pumps for reabsorption. It therefore also serves as a Ca store or reservoir. As the myosin heads do not synchronise with each other and therefore there are always enough myosin heads bound to the actin, the myosin filament cannot slip back on the actin. The neurotransmitter that transmits the nerve impulse for contraction to the muscle is acetylcholine. A concentric contraction therefore results from the fact that the myosin and actin filaments continue to interlock through repeated cross-bridge cycles and pull the Z-discs towards the M-discs. One cross-bridge cycle produces approx. 1% of the possible contraction of the muscle. As calcium and magnesium are involved in the mechanism, an imbalance of these electrolytes can have a direct influence on the performance of the muscle and may also cause cramps (Ca deficiency).
Titin
Titin (connectin) filaments run through the sarcomere from the Z-discs to the myosin filaments, keeping them in position parallel to the actin proteins. Titin contains 10% elastic areas that make the muscle stretchable. It is not involved in contraction itself, but determines elasticity and contraction speed and prevents an unphysiological increase in muscle length. At 36 megaDaltons (1 Dalton is 1/12 of the mass of a C atom and therefore roughly the mass of a nucleon), it is the largest known human protein. At the edges of the sarcomeres is an area that is not reached by myosin.
The area around the M-disc that does not come into contact with actin is known as the H-zone (Hensen’s zone). The I band/zone (isotropic, single refractive index) is the band around the Z disc that only contains actin and titin. The remaining area is referred to as the A band (anisotropic).
The discs and zones are therefore arranged as follows:
Z – I – A – H – M – H – A – I – Z . Put simply, contraction takes place by docking myosin to actin, whereby the angle of the head of the myosin filament changes, resulting in a movement of the myosin relative to the actin.
The contraction is triggered by a nerve impulse. Calcium and magnesium are involved in the mechanism, so that an imbalance of these electrolytes has a direct influence on the performance of the muscle and may cause cramps. Furthermore, ATP is required, which is split into ADP and a phosphate residue and must be replaced. Such a „cross-bridge cycle“ lasts approx. 10 – 100 ms and shortens a sarcomere by 0.1-0.2 µm, but the cycle can be repeated several times, whereby the muscle or the individual sarcomere can be shortened by approx. 50% within one second by about 50 cycles.
Sarcomere length
the sarcomere length is the actual length of the sarcomere, i.e. the distance between two Z-discs and therefore a measure of how far a muscle can still contract or how far it can still stretch. The average resting length of a sarcomere is around 2 – 2.5 µm, at just over 1 µm the muscle is fully contracted and no further contraction or shortening of the muscle is possible. The maximum length of a muscle is reached at just over 4 µm.
bilateral strength deficit
The bilateral strength deficit describes the fact that when performing identical activities bilaterally, the muscles involved provide less strength than when the same movement/posture is performed unilaterally; the simultaneous contraction force of both limbs together is therefore less than the sum of the individual forces of the right and left sides. a40> of both limbs together is therefore less than the sum of the individual forces of the right and left sides. The background to this phenomenon lies in the reduced neurological control of the muscles during bilateral activity. The exact causes and mechanisms are not yet fully understood, even though studies have sufficiently proven the fact itself. For training, this can mean that exercises performed unilaterally (in continuous side changes or in side changes after a set) enable more effective strengthening. Unilateral execution also allows for better concentration on the respective agonists and more precise execution. However, better concentration on the respective agonists does not seem to be the predominant reason for the bilateral strength deficit. If the exercise is performed with continuous side changes, the recovery time between the individual repetitions on the same side is significantly longer than if the side change only takes place after a set, which also leads to greater performance and presumably greater training success. Another effect that could already take effect with slow execution speaks in favour of continuous side changes: muscle contraction can impair intramuscular supply, so a relaxation phase of the muscle, even if short, improves this. Another advantage of unilateral execution is that other parts of the musculoskeletal system that have a supporting function are significantly less prone to overload than with double external load. This is often seen in the lumbar region during bilateral squats. In addition to the higher maximum strength achieved with unilateral execution, these exercises also have the advantage that much more stabilising muscle tissue in the kinetic chain is used and trained. In the split squat, for example, the lower leg muscles are required to a much greater extent and also more intensively, especially in terms of the restraint system of the ankle joint, which plays hardly any role in bilateral squats. Of course, this also applies to the hip muscles, albeit to a greater extent for the ankle joint, as the foot, with its very narrow width, provides an extremely narrow physical support base. Bilateral strength deficits therefore primarily affect bilateral sports such as rowing and squats. Not least because of this, single-leg exercises such as split squats have become increasingly popular in fitness and professional sports. The phenomenon of bilateral strength deficit should not be confused with general strength deficit, in which the maximum voluntary strength falls significantly short of the theoretically possible.
general strength deficit
The general strength deficit describes the fact that the maximum voluntary strength is significantly lower than the theoretically possible strength. This means that it is not possible to voluntarily utilise the potential maximum strength, possibly because the body wants to protect the musculoskeletal system, especially the muscles and their tendons, from overload. Another reason could be that the body does not want to use up all its reserves in a single movement so that it is still possible to perform another movement if necessary. In exceptional psychological situations, however, the potential maximum strength can be provided under certain circumstances.
Forms of contraction
When contracting a muscle, a distinction is made between
- isotonic: the muscle shortens without changing the force, as is the case when lifting an object very slowly
- isometric: (see here) The length of the muscle remains constant, regardless of the force exerted. Examples: Trying to lift an object that is too heavy or trying to open a locked door
- eccentric: (see here)
- concentric: (see here)
- auxotonic: both force and length change.
The myosin filaments have two heads. During concentric contraction, only one of them binds at a time; during eccentric contraction, both also bind, which explains why a muscle has significantly more force during eccentric contraction.
Heads and bellies
A muscle can not only have several heads such as the biceps or triceps brachii, but also several bellies such as the rectus abdominis. The contractile abdomens are separated from each other by non-contractile tendon tissue (insertions).
The type of fibre
A distinction is made between red fibres (slow-twitch, type 1) and white fibres (fast-twitch, type 2) in the muscle fibres of the skeletal muscles.
Typ 1
The red musculature has more muscle fibres with oxidative energy production and is therefore well suited to endurance performance, but can develop significantly less strength.
Typ 2
The white one is better suited to high-speed strength and power performance, but can convert less energy continuously in real time and therefore tires more quickly.
The ratio of the types differs depending on the muscle and between individuals. It can be influenced to a limited extent by training. People with a higher proportion of white fibres respond better to training stimuli, their muscles are more visible, but their muscles become exhausted more quickly.
The soleus is one of the muscles with a high proportion of type 1 fibres; depending on the source, this is given as 60-80%.
The tibialis anterior also has a high proportion of type 1 fibres, whereas vastus lateralis pars quadriceps and biceps brachii are reported to have a maximum proportion of 50%, often less. Triceps and deltoids also have low type 1 fibre proportions, gluteus maximus and the autochthonous back muscles have higher type 1 fibre proportions.
The fibre composition of a muscle is genetically predisposed.
Basically, trunk muscles are type 1-rich, while the muscles of the limb arm, for example, are mainly type 2a and 2x.
Through strength training, mainly type 2a and type 2x fibres hypertrophy and type 2x partly convert to type 2a.
Type 1 fibres hardly hypertrophy at all, but do become stronger.
Transformation from both sides is known from sprint training:
both type 1 and type 2x partly convert to type 2a.
The myokine BDNF is involved in the conversion of type 1 fibres into type 2 fibres. It can be shown that strength training actually impairs endurance performance to a certain extent (muscularly).
Endurance training, on the other hand, causes improved capillarisation and proliferation of mitochondria (mitochondrial biogenesis) in all three types (to varying degrees). In addition, there is a conversion from type FF (2x) to type FR (2a) and type FR (2a) to type ST (1a). In fact, the innervation of type 1 and type 2 differs in such a way that type 2 is innervated by faster nerve pathways. In top cross-country skiers, for example, a change in type 2 fibres towards type 1 has been demonstrated, such that they achieve a similar capillarisation and a similar mitochondrial volume. In strength training, a distinction must be made between high-speed strength training, which shifts from type 1 to type 2 and acquires type 2x in fast movements, and strength endurance and endurance training, which shifts from type 2a towards type 1a. The changeability of the fibre profile is referred to as plasticity. Prolonged intensive training and longer training breaks have a particularly plastic effect.
Contrary to expectations, ageing and immobilisation shift the fibre profile from type 1a towards 2a and 2x. However, as the maintenance stimuli in these situations are low-threshold or absent, strength is reduced, which also leads to reduced stability in addition to the loss of strength that is to be expected anyway.
Feathering
Pinnation refers to the property of muscles that their fibres do not run parallel to the longitudinal extension of the muscle. With single pinnation (unipennatus) the deviation is uniform, with multiple pinnation it occurs in different ways. The physiological cross-section (section orthogonal to the course of the fibres) increases in comparison to the anatomical cross-section (orthogonal section through the macroscopic course of the muscle) as a result of pinnation. Fibrillation therefore means that the direction of the fibre course is different from the connecting line between the origin and insertion, which means that a larger number of muscle fibres can attach to the tendons with the same muscle thickness (anatomical cross-section), which increases the force transmitted to the tendons (tendon force) and also increases the stroke height of the muscle. The angle of the pinnation changes slightly with the contraction.
Example: if a muscle is pinned at 45°, it loses a factor of cos (45°), i.e. around 30% of its force due to the changed direction of the muscle fibres, but the physiological cross-section increases by a factor of 3, so that the gain in force that is transferred to the tendons is a factor of 2.1. The average force of around 40 N / cm² has therefore increased to 84 N / cm² diameter. With the greater force, the maximum force and thus the maximum contraction speed naturally also increases.
Contraction
The contraction of a muscle or muscle contraction refers to the process in which a muscle applies contraction force beyond its resting innervation by consuming more energy (compared to the resting state) in the form of hydrolysis of ATP to ADP. A chemical reaction of the protein myosin, which is attached to the centre disc of the sarcomere, causes a geometric change in its head, which docks onto variable sites of the protein actin, which is attached to the Z-disc bounding the sarcomere, leading to a shortening of the sarcomere.
The process from one force impact to the next is known as the cross-bridge cycle. A distinction is made between concentric, eccentric, isometric and isotonic contraction. If nothing else is specified, contraction is generally understood to mean concentric contraction. For a more detailed description, see the structure of the sarcomere and the description of the contraction mechanism on Wikipedia.
Tonic muscles
Tonic muscles are those that primarily fulfil a holding function and are used less for rapid movement. As a rule, they tend to be of fibre type 1 and tend to shorten and harden with prolonged holding work, lack of exercise and excessive strain. With tonic muscles, it is particularly important to avoid a chronic increase in tonus and their shortening.
Phasic muscles
In contrast to tonic muscles, phasic muscles, which are primarily used for movement and are predominantly of fibre type 2, i.e. white, must above all be strengthened or kept strong. There must be no imbalance between the two groups of muscles, as can easily occur with prolonged identical postures and chronic lack of exercise.
Forms of contraction
Eccentric contraction
Muscle contraction, in which the interlocking of actin and myosin in the sarcomeres decreases, i.e. the distance between the M and Z discs increases, so that the distance between the origin and insertion of the muscle increases. In addition to the eccentric contraction, there is also the concentric, the isometric and the isotonic contraction. The eccentric contraction is also known as pliometric contraction (plio: gr.: long). The properties of the eccentric contraction known today are
- Increased power development
- Structural additional load on the activated muscle fibres
- favourable energy balance (low cost, high force)
- Influence on the voluntary innervation, altered neuromuscular innervation pattern
- Tendency to overuse-related injuries to the muscles
- Increased risk of articular and periarticular injuries and tendons
- Effective form of training to counteract sarcopenia and reduce the risk of osteoporosis
- Favourable in the treatment of tendopathies and for the prevention of muscle injuries
Walker et al (7) showed in a 10-week study that eccentric training in particular (concentric movements were massively facilitated for the test subjects) with 40% higher weights showed a comparable increase in muscle mass compared to classic strength training, but was superior in the increase in maximal strength.
isometric contraction
Muscle contraction, in which the position of actin and myosin in the sarcomeres and the distance between the M and Z discs remain unchanged and therefore the distance between the origin and insertion of the muscle also remains the same. For forms of training with an isometric component, it should be borne in mind that with isometric movements from 10% of the maximum force, the effect can be demonstrated that the increased internal muscle pressure (the fasciae are hardly stretchable) reduces the intramuscular supply. From 60%, the pressure is so high that the intramuscular supply is completely interrupted and only anaerobic metabolism takes place intramuscularly. This not only causes metabolic stress on the arterial supply side, but also on the opposite side, the removal side, because if the smallest arterial vessels are compressed to the point of impermeability, the venous vessels are even more so. Metabolic end products such as lactate, inorganic phosphate and H+ accumulate. This in turn leads to an increased release of testosterone, growth hormone and the insulin-like growth hormone IGF 1.
Isometric contraction is particularly interesting for tendons:
- It increases rigidity, thereby increasing energy storage and release.
- Isometric training can be very gentle on damaged joints, thus facilitating rehabilitative training.
- It is very suitable for the rehabilitation of tendinopathies.
- It promotes collagen synthesis.
- Isometric training often has a pain-relieving effect in cases of tendinopathy. In therapeutic application for tendon disorders, a holding time of 45 seconds is recommended, if possible with a load of up to 90% of maximum strength and 5 sets (study by Rio et al.). 70% is considered the minimum load for promoting tendon growth. However, the study situation on this topic is currently unclear.
In addition to the isometric contraction, there is also the eccentric, the concentric and the isotonic contraction.
isotonic contraction
Muscle contraction in which the tension of the muscles (contraction force) remains the same. Whether an eccentric or concentric contraction results, i.e. whether and how the distance between the origin and insertion of the muscle changes, depends on the external resistance to the contraction. This includes not only forces acting on the body from the outside, but also the tension of the antagonist(s). In addition to the isotonic contraction, there is also the concentric, the eccentric and the isometric contraction.
concentric contraction
Muscle contraction, in which actin and myosin in the sarcomeres interlock further, reducing the distance between the M- and Z-discs, so that the distance between the origin and insertion of the muscle is reduced. In addition to the concentric contraction, there is also the eccentric, the isometric and the isotonic contraction. The concentric contraction is also known as miometric contraction (mio: gr.: short). With concentric contraction against a certain resistance, a muscle develops less force than with eccentric contraction. This dependence is also dependent on velocity, which was described by Hill in the equation of the same name, which states that velocity is inversely related to force, see force-velocity function.
Function reversal
Function reversal is the ability of a muscle in a certain joint to support a different movement in a certain angular range of a movement dimension or direction than in another angular range. This does not imply that this muscle may also perform a functional reversal in other movement dimensions. The reversal of function can be derived purely geometrically, as a muscle always shortens the distance between the insertion and the origin through its concentric contraction. The two best-known examples are the tensor faciae latae, which supports flexion in the knee joint via the tractus iliotibialis below 30 or 40 degrees, depending on the literature, extension of the knee joint, but then the flexion of the knee joint. If we consider the iliotibial tract as a kind of extended tendon of the tensor faciae latae, the reversal of function therefore takes place in the second joint it covers. An example of a monoarticular muscle with a reversal of function is the pelvitrocrural piriformis, which supports up to 60 degrees of flexion in the hip joint to reduce flexion or extension and exorotation in the hip joint, from 60 degrees flexion but further flexion and endorotation in the hip joint.
Another example of monoarticular muscles with functional reversal are the adductor muscles, almost all of which, with the exception of the ischiocondylar portion of the adductor magnus, from small flexion lines of the hip joint support the flexion, but from very wide flexion lines support the reduction of the flexion. This in turn can be explained purely geometrically: in zero flexion in the hip joint, the attachment is located far inferior to the origin, but in very wide flexion of the hip joint it is very far cranial to the origin. In between, the adductors assume a minimal length with the same degree of abduction or adduction.
In the same way, the pectoralis major has a functional reversal in that it pulls the arm forwards from wide retroversion due to its ventral origin, but also adducts the front again from wide frontal abductions. Theoretically, it should also be possible to find a large degree of retroversion in very mobile people, in which the latissimus dorsi has a retroversion-reducing effect, but at best up to an angle of around 10 or 20 degrees retroversion, after which it has a frontal adducting and slightly retroverting effect. The phenomenon of functional inversion has nothing to do with Lombard’s phenomenon, as it occurs in the ischiocrural muscles in that they appear to support the stretching of the knee joint in a closed kinetic chain during walking or running. This results from the fact that they are also extensors of the hip joint and produce an effect there due to the larger lever arm in the hip joint, which, due to the closed kinetic chain with the foot as punctum fixum, supports the extension of the knee joint, although they nonetheless generate a flexing moment in the knee joint, but this is outweighed by the movement of the thigh in the hip joint is outweighed .
The deltoid also has a functional reversal, in particular the spinal and clavicular pars, which run up to an angle of lateral abduction of around 60° in their connection from origin to insertion mediocaudally of the centre of rotation in the shoulder joint, so that they cause a lateral adducting moment, and only from this angle of lateral abduction craniolateral to it, so that they can have a lateral abducting can act .
Depending on the exact position of the Tuberositas radii, the biceps brachii also has a functional reversal, but this is in very wide supination, so that it has a supination-reducing effect from the last degrees of supination, i.e. pronating, but it has a supinating effect over the widest range of supination, and this becomes stronger the further the elbow joint moves towards 90°.
A reversal of function is therefore always given when the connecting line between the attachment and the origin changes its position in relation to the centre of rotation in such a way that the centre of rotation can change to the other side of the straight line through the attachment and origin, i.e. the triangle formed from the three points changes its orientation from clockwise to anti-clockwise or vice versa. This applies provided that the muscle runs in a straight line and its force is not diverted. Therefore, it does not join the deltoid pars acromialis, as its force is diverted via the humeral head, unlike the other two parts.
Naturally, the torques that can be achieved in the joint continue to decrease towards zero from both sides of a muscle with a functional reversal to the angle of the functional reversal. The maximum torque is therefore far away from the point of function reversal usually at one and the other end of the movement space.
Contraction force
The force with which a muscle contracts, i.e. pulls its insertion and origin towards each other. The contraction force is most simply understood as the axial tensile load applied in one (some muscles of the movement apparatus do not attach at both ends with tendons, but possibly directly to bones) or both tendons, the tendon force.
The contraction force of a muscle depends on how many motor units are activated and how many muscle fibres (ranging from approx. 100 to 3000) they comprise. A motor unit comprises a single motoneuron together with all the muscle fibres innervated by it. The motor units with few muscle fibres allow a fine gradation of force (muscles of the eyes and some of the fingers), those with many fibres belong to muscles that can exert a lot of force, e.g. quadriceps. During a muscle contraction of increasing strength, first small motor units are used (recruited), then larger ones (Hennemann’s principle). The fibres of individual motor units do not lie next to each other but are distributed throughout the muscle. For maximum force development, the neuronal impulse frequency is increased so that individual contractions overlap and their force is totalised.
If the movement system is given a task, all relevant muscles must provide a certain force, the motor force. The maximum achievable motor force is referred to as maximum force and depends on various influencing variables, including psychophysical factors such as motivation and inhibiting variables such as fear.
Maximum force
The maximum force is the greatest force a muscle can provide voluntarily. On the one hand, this can be determined as isometric maximum force, so that any greater force isometrically can no longer be maintained, and on the other hand, and very frequently used in application, as 1RM, i.e. as single-repetition maximum force. In a repetition of a cyclic movement, the concentric contraction is always the greater requirement, so this is relevant here. Basically, the isometric maximum force depends on the current
sarcomere length and the concentric maximum force depends on the exact working range (the sarcomere lengths traversed) and the execution speed. The greater this is, the lower the maximum force that can be achieved.
In the case of maximum strength, protective mechanisms take effect that prevent all the strength potentially available from the muscle from being mobilised by autonomously holding back reserves that are only activated in physical-pathological situations or exceptional psychological situations, so that the so-called absolute strength is then available. The difference between absolute strength and maximum strength is known as the strength deficit. The strength deficit can be estimated by the number of repetitions that can be performed with 90% of the 1RM. On average, this is 3-5 repetitions. If there are more, neuromuscular activation (intramuscular coordination) is poor and the strength deficit is above average. The strength deficit can be trained.
Apart from these factors, there are essentially three areas that influence the achievable force:
- neuronal influences:
- Recruitment: how many motor units can be recruited at the same time?
- Frequency: how quickly can the nerve impulses stimulate the muscles to contract (number of action potentials: motor units from ST fibres are innervated at up to 20 Hz, motor units with FT fibres at up to 50 Hz).
- Synchronisation of the motoneurons: increase in simultaneously active motoneurons
- Inhibitionsabbau
- Reflex support
- muscular influences:
- Muscle cross-section (hypertrophy)
- Muscle fibre composition (distribution of fibre types, see below)
- Muscle fibre characteristics
- Metabolic quality
- Capillarisation
- Muscle elasticity
- anthropometric or biomechanical conditions:
- Leverage ratios, e.g. as force arm: distance chord-centre of rotation
- Joint condition
- Vision elasticity
- external conditions that may play a role, such as temperature, time of day, surface and footwear
In addition to the absolute muscle strength defined above, relative muscle strength is often relevant, i.e. the absolute muscle strength in relation to body mass.
Force-length function (muscle-force curve)
The isometrically achievable muscle strength depends on the possible number of cross-bridges between actin and myosin. This in turn depends on the current sarcomere length. Below around 1.27 µm and above around 3.65 µm, the muscle can no longer generate any significant force. The functional dependence of the maximum isometric force on the length of the sarcomere is roughly described by an upside-down parabola that is slightly elongated in the direction of greater lengths. In reality, however, it is polygonal in small steps because the increase and decrease of the force depends linearly on the integer number of bridges that can be reached.
Starting at the position of maximum elongation, more and more myosin heads can engage with the actin as the contraction increases and contribute force with their geometric change. Shortly after reaching the maximum contraction force, the opposite effect takes effect: the binding sites on the actin to which the myosin heads can dock become fewer and fewer until finally actin and myosin have maximally interlocked, no further docking sites on the actin follow, and further contraction is no longer possible. At this point, the contraction force has dropped to zero. There is also a zero point of the function on the other side of the curve (furthest elongation), namely where no myosin head has yet intervened in the actin (elongation, not in monoarticular muscles).
The maximum force is reached at an average sarcomere length of around 2.8 µm. If, in addition to the active force generated by contraction, the passive force that an „averagely stretched“ muscle exerts towards the end of its maximum sarcomere length against further stretching is also considered, this curve typically increases exponentially towards 4 µm after a local minimum (depending on the individual state of stretching).

Force-velocity function („Relation„)
The maximum muscle power (remember: power = work / time = force * speed) is the product of shortening speed and achievable muscle force. The achievable muscle power remains largely the same across all possible speeds, a slow concentric contraction enables greater force than a fast one, isometrically more force is available than concentrically and eccentrically even more.
The force-speed function (incorrectly also -relation) describes the force that a muscle can produce under comparable conditions (e.g. with regard to the status of regeneration, activation/preactivation) at different contraction speeds. This is described by a curve with an inflection point during isometric contraction. It runs in the second quadrant (negative speed means eccentric contraction) approximately asymptotically against the eccentric maximum force, has an inflection point on the Y-axis (zero speed means isometric contraction) in the isometric maximum force and runs left-curved against the x-axis with a zero point at maximum contraction speed.
At maximum speed, the achievable force is zero, the fastest possible eccentric contraction enables maximum force – with maximum risk of injury.
Muscle training takes place mainly, but not only, in the right part of the curve and enables an increase in muscle performance. Training can be roughly divided into maximal strength training and speed strength training. If the axes in the Hill diagram (Hill, 1938) are swapped, this results in an increase in leg A due to maximal strength training (training with >= 90% of maximum strength) and an increase in leg B due to high-speed strength training. This means that in the case of gravity, a higher load can be moved at the same speed and a higher speed can be achieved with the same load. In the case of high-speed force, this means that a higher load can be moved at the same speed or a higher speed can be achieved with the same load. Incidentally, the Hill model is only valid under strong conditions, above all only when isometrically considered, which is why it is replaced by the Häufle model, which provides a viscoelastic (shock absorber-like) element parallel to the contractile element and depicts real measurement data incomparably better.
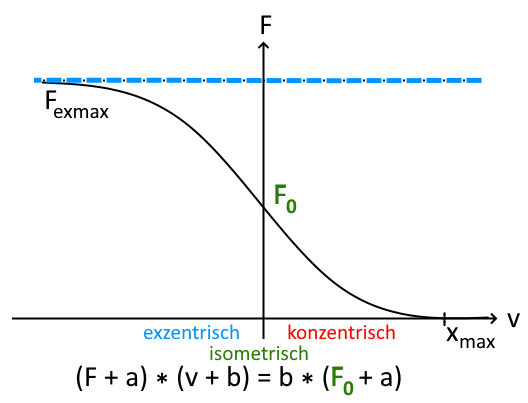
with v as the contraction speed, F as the muscle force, F0 as the isometric maximum force and empirical constants a,b.
Simplified, this means that the faster a muscle contracts, the lower its force and the slower the eccentric movement, the greater it is.
Strength endurance
Strength endurance usually refers to the ability of the body or a part of it to exert a greater force of at least 30% of the maximum force (in terms of 1RM) dynamically or isometrically over a longer period of time. Definitions of strength endurance also often refer to resistance to fatigue during such exertion. Since strength endurance must always refer to specific muscles, the term local anaerobic dynamic or static endurance is more appropriate, whereby in common considerations the size of the muscles used in practical application in sports is usually given as roughly one fifth of the body’s muscle mass. This term therefore does not apply in the required generality and does not apply everywhere where it would make sense to speak of strength endurance, as the following examples show.
Isometric case
If, for example, the force of the finger flexors were required to hold a fist closed with a required minimum force that is not too low, for example to hold a very heavy object for 2 minutes or to compress an object, the relevant muscle mass of the finger flexors of one hand would be far below this limit. However, this would be a good example of a narrowly localised isometric strength endurance performance. A person with the same maximum strength in the finger flexors, who is not adapted to this requirement, would run into localised overacidification of the performing muscles much faster than someone specially trained for this.
The holding power from the example becomes anaerobic if it is above a non-uniformly specified limit of around 20% of the maximal force of the relevant muscles in the sense of 1RM, as isometric contractions of this magnitude cause compression of the vessels in the muscle, which impairs the supply to such an extent that the aerobic metabolism is impaired. It then depends on how well the muscle can perform anaerobically, which depends heavily on the energy stores and the enzymes stored. This factor can be trained. Below 10-15% it can be assumed that the supply is still sufficiently good for a largely anaerobic metabolism, while at 50-60% the intramuscular supply comes to a complete standstill.
Another case is carrying heavy objects with bent arms, various heavy holding activities or intensively performed asanas against corresponding internal or external body resistance such as gravity. In sport, holding power is less frequently required. One example would be the downhill squat in ski racing, where dynamic load peaks and unloading can occur due to uneven terrain, another would be the holding work in climbing.
Dynamic case
In the dynamic case, there are usually cyclical movements in which the TUT exceeds the recovery time by a minimum factor, and the individual repetition is heavy enough so that a continuous aerobic performance is not possible for the muscles involved in average untrained people, because the energy consumption of the muscles and tissue compression are no longer in a favourable ratio for an aerobic metabolism. As cyclical movements can, but do not have to, include recovery phases, the analysis here is much less simple than in the static case with the limit of around 20% of the 1RM. For example, a bicep curl can be performed as a clearly cyclic movement completely without relief and recovery for the arm flexors, whereas when cycling without a click or bar, the quadriceps are only used during the extension movement to push down on the pedal and have the other half of the cycle to recover.
Dynamic strength endurance requirements are also often found in manual trades, for example for the finger flexors, when an electrician continuously cuts through countless cables with pliers while wiring a control cabinet or a fitter continuously screws or unscrews things with countless arm movements while exerting additional force on his finger flexors.
Classic examples of sports in which strength endurance is required in its dynamic form are cycling or rowing, in which the individual movements are sufficiently difficult.
In dynamic training to promote strength endurance, sets of 15 – 30 repetitions are typically completed at 40-60% of the 1RM, although this is very vague because the speed of execution plays an important role and must be specified. A more precise definition is via the TUT, which should be 50 to 120 seconds per set for this type of training. 30% of 1RM is usually regarded as the lower load limit for strength endurance training.
Strength endurance capacity is therefore not a pure combination of endurance capacity and high maximum strength, even if good endurance capacity of the organism and high maximum strength of the muscles concerned are important. Rather, it is a specific ability not only of the organism as a whole, but above all of the muscles concerned, which must be specially trained for this purpose. The specific training for this must be aimed above all at the vascularisation of the muscles. Accordingly, practice shows that people who perform strength training with averagely fast movements and low repetition rates, so that they achieve a considerable maximum strength, and also perform an endurance sport such as running or cycling, may not necessarily be able to achieve a good strength endurance performance with the same muscle groups that they train in strength sports.
The physiological adaptations to the strength endurance requirement are primarily improvement of the buffer capacity, oxidation capacity, improvement of the vascularisation (capillarisation) of the muscles and the mitochondrial density in the muscles, increase in the metabolic activity of relevant enzymes and sarcoplasmic hypertrophy.
In practical application, strength endurance is tested either isometrically up to isometric muscle failure with a suitable proportion of 1RM or as the maximum achievable number of repetitions up to concentric muscle failure in a cyclic movement with a suitable proportion of 1RM and a predetermined execution speed. A distinction is sometimes made between „aerobic strength endurance“ with loads of 30-50%, „medium strength endurance“ with loads of 50-75% and „high-intensity strength endurance“ with loads of up to 100%.
Predeexpansion
Pre-stretching refers to the muscle length (more precisely: sarcomere length) at the start of contraction. For good force development, myosin heads must have sufficient connection to an actin filament, which is the case below about 2 – 2.2 µm. From around 3.6 µm, actin and myosin no longer have a connection, so in order to enable contraction, synergists of this muscle must change the joint positions appropriately. Mathematically, this results in a maximum possible contraction of the muscle of around 60%, but in practice this is more like 30%.
Articulation
monoarticular
Single-jointedness of a muscle, i.e. the property of a muscle to span exactly one joint and cause a movement in this joint. In general, a muscle of the movement apparatus or its tendon causes flexion of the joint on the inside of which it runs.
biarticular
Two-jointedness of a muscle, i.e. the property of a muscle to span exactly two joints and cause a movement in these. The movement can be the same in both joints (flexion or extension), such as in finger joints or the spine, or also flexion in one joint and extension in the neighbouring joint (example: M. rectus femoris). However, this is more due to the nomenclature: the rectus femoris pulls the lower leg towards the thigh ventrally, just as it pulls the thigh towards the pelvis ventrally. In this sense, it is a rectilinear movement. However, the movement possibilities and habits of humans suggest that the movement of the lower leg dorsally in relation to the thigh should be described as flexion in the same way as the movement of the thigh in relation to the pelvis ventrally.
However, there is also a case in which a muscle causes geometrically opposing movements. The lumbricales flex the metacarpophalangeal joints (MCP) and extend the proximal phalangeal joints (PIP). This is possible by switching the executing tendons to the other side: from palmar in the MCP to dorsal in the PIP.
In addition to biarticular muscles, there are also monoarticular (covering one joint) and polyarticular (covering more than two joints).
polyarticular
Multi-articularity of a muscle, i.e. the ability of a muscle to span more than two joints and cause movement in them.
Muscle spindle
Proprioceptors with differential (measure the change in length) and proportional properties (measure the length), which report the stretch state of the muscles to the CNS. Ultimately, the elongation state is essentially equivalent to the sarcomere length. If the muscle is stretched quickly, a muscle stretch reflex (an intrinsic reflex) is triggered, which sends a contraction stimulus to the same muscle via the alpha-motoneuron. Renshaw cells limit this stimulus by means of negative feedback, so that a very strong innervation is attenuated and good fine motor skills can be imparted, especially to the hands and feet, which lie far distally. One of the best-known examples is probably the patellar tendon reflex.
Training
The effects you are aiming for during training include
- Hypertrophy: Enlargement of the muscle fibre (thickness growth, increased number of parallel sarcomeres)
- Hyperplasia: Increase in muscle fibres, the ability to do this in humans is controversial
- longitudinal adaptation (see also below): depending on the ROM used, the serial number of sarcomeres can be reduced (e.g. immobilisation at a rather small distance origin-approach) or increased (especially through training or active contraction beyond everyday stresses with large ROM). The increase in serial sarcomeres increases the muscle performance and thus the contraction force and the unloaded maximum shortening velocity, even without radial hypertrophy (increase in cross-sectional area). The first results on this were obtained by Lynn and Morgan in 1994 in experiments with the vastus intermedius of rats. Vigorous eccentric exercise in large sarcomere lengths leads to damage to the sarcomeres, which can apparently also be repaired with an increase in serial sarcomeres. This shifts the working range of the individual sarcomeres somewhat and their stability gains somewhat, which means somewhat reduced susceptibility to injury.
Incentives for muscle building
There are three important factors for muscle building:
- Training with growth stimuli (heavy weight with few repetitions)
- Regeneration: the electrolytes lost through sweating are quickly replaced, the replenishment of glycogen stores takes a little longer and benefits from the intake of KH. This part of regeneration takes hours to days and requires a constant supply of protein, ideally starting no later than 20 minutes after training. The resynthesis of mitochondrial proteins takes longer than the replenishment of glycogen stores. Regeneration depends heavily on the intensity of sleep and nutrition.
- Nutrition
In 2010, Schoenfeld concretised the growth stimuli in relation to the muscle itself as:
- Mechanical tension
- metabolic stress
- Muscle damage (microtrauma)
However, muscle damage did not prove to be a sustainable factor (Schoenfeld 2012; Damas, Libardi & Ugrinowitsch, 2018). In 2016, Schoenfeld, Ogborn & Krieger even postulated that training that causes a high degree of muscle damage leads to longer necessary regeneration with reduced resilience, which is no longer available as time for high-intensity training, so that the overall training success is then lower.
The extent to which metabolic stress contributes directly to training success has not yet been conclusively clarified. However, an indirect contribution can be considered certain, as the underlying undersupply forces a greater degree of muscle fibre recruitment and thus increases the mechanical tension across the entire muscle (de Freitas, Gerosa-Neto, Zanchi, Lira & Rossi, 2017; Schoenfeld et al., 2017). For maximum hypertrophy, both maximum recruitment of muscle fibres and maximum (or maximum tolerable) maximum tension (contraction force) must be achieved. Recruitment is not only dependent on the load, but also significantly on the contraction speed, the slower the more. Therefore, the factor TUT and also the relation of TUT to the regeneration time within the load are of great importance. It should be noted that physiologically, with increasing contraction speed, fewer and fewer cross-bridges can be formed at the same time.
With increasing peripheral fatigue, the excitation thresholds of the large motor units are lower, so that complete fibre recruitment can be achieved even with significantly submaximal resistances (e.g. weights). This can already be achieved at 30% of the maximum load (1-RM). In other words, a certain approximation to muscle failure (peripheral fatigue) is necessary and sufficient for a strength training-induced increase in protein synthesis in the muscle (Schoenfeld, Grgic, Ogborn and Krieger, 2017).
One training session per week is sufficient for maintenance training. In addition to the familiar forms of training, which involve constant, increasing or decreasing weights per training session, a study has shown that wave-shaped training, a split training programme in which each muscle group is trained three times a week with different intensities and repetitions, each once with 85%, 70% and 55% of the maximum strength, is promising.
The number of repetitions plays a much smaller role than the intensity and TUT (time under tension). Isometric contractions enable large increases in strength in a short time, but are not suitable as the sole training method. The resulting high metabolic stress is an important hypertrophy stimulus. Reducing the outflow and inflow of blood can also increase the hypertrophy stimulus (KAATSU training, in which a blood pressure cuff compresses the tissue and therefore the blood vessels.)
A distinction is made between the duration of the training stimulus according to TUT:
< 20 s: Maximum strength
20 – 50 s: Hypertrophy range
> 50 s: strength endurance
Jo Weider, who has been involved in strength training and bodybuilding all his life and has worked with many athletes in this field, and was partly responsible for the success of Arnold Schwarzenegger, among others, has created a collection of „training principles“ which, in his experience, summarise the most important principles for stimuli for hypertrophy and strength gain, see here.
Trainingsparameter
In physical activities, whether in sport or everyday life, a distinction can be made according to the intensity of the movements or postures into
- Low-threshold, i.e. those that can hardly provide a maintenance stimulus for the muscles or the circulatory system
- Medium-threshold, i.e. those that do
- supra-threshold, i.e. those that stimulate the body to adapt in the sense of better performance
Suprathreshold stimuli are characterised by a more or less pronounced proximity to muscle failure, for which the greater the level of the load or more the duration of the demand can be used. A simple example from the field of strength training is used to show that there are some free parameters that influence the body’s adaptation to a given level of a suprathreshold external load. Movement takes place in joints and is usually gravity-induced, otherwise externally induced or muscle-induced. For this example, we will look at a simple muscle-induced movement, the bicep curl. As with many other movements of parts of the extremities, it is not possible to train a single monoarticular muscle, as the name of the exercise suggests, but rather the monoarticular brachialis, the biarticular biceps brachii, the brachioradialis and, very subordinately, some forearm muscles with origin at the condyles of the humerus. If we keep the shoulder joint absolutely still for this analysis, the biceps brachii behaves a bit like a monoarticular muscle. Of course, this is not entirely true: the vertical upper arm of the biceps curl shifts the working range towards greater sarcomere lengths compared to a widely raised arm. This is very favourable for training, as it is known that growth stimuli can be set more easily in longer sarcomere lengths. The best-known free parameters that can be varied include the duration of a single repetition and the number of repetitions. In addition, similar to the basic principles of pranayama, a holding phase could be inserted at the end of the concentric contraction or at the end of the eccentric contraction. The exact speed of the movement is also relevant. For example, it makes a difference whether a large proportion of the time is spent in greater sarcomere length, which provides a greater growth stimulus, but also better maintains or even increases mobility. This becomes clear as the limit value of this parameter would be an arbitrarily long holding time at maximum sarcomere length, which would presumably even lead to longitudinal muscle adaptation and therefore clearly promote mobility. If instead a large part of the time was spent in short sarcomere length, this exercise would reduce the mobility of the muscles. Here, too, the limit value formation makes this clear: a long hold in the shortest sarcomere length causes an adaptation in this direction up to negative longitudinal muscle adaptation. The graph schematically depicts a time course in which moderately pronounced, less time is spent in shorter sarcomere lengths and more time in longer ones. This is shown by the reciprocal curves of contraction speed and flowing time average in a sarcomere length. Again in analogy to pranayama exercises, holding phases can also be introduced, which can significantly extend the total duration of a repetition and represents a further parameter that can be used to approach muscle failure.
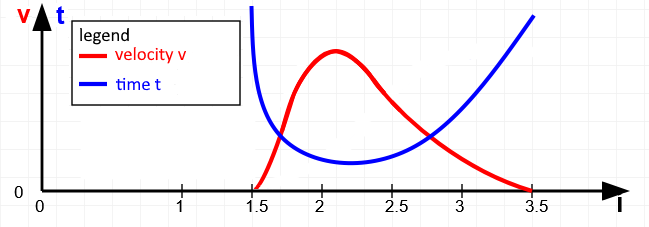
The better-known training parameters certainly include the number of sets, the weight increase within the sets and the number of repetitions, which can inevitably be limited by the selected weight. However, the pauses between the sentences are also a not insignificant parameter. If the pause is only a few seconds, it can be neglected as such. It is then as if two sentences were strung together to form one long one. If the pause is significantly longer than 3 minutes, it can be assumed that the muscle’s willingness to perform has already begun to decline. There are different opinions and recommendations on this subject. Another interesting aspect is supersets, which under favourable conditions represent an almost continuous challenge for the energy supply. Usually, two muscle groups are trained in alternation with each other, whose kinetic chains hardly overlap, for example bench press for the pectoralis major, which includes the stretcher of the elbow joint, alternating with lat pulls or rowing, that instead of the extensors of the elbow joint its flexors are also trained and mainly on the dorsal shoulder joint adductors such as latissimus dorsi, Teres major and Teres minor.
Supercompensation theory
After completing regeneration from heavier training or intensive physical work of a non-sporting nature, the body ensures a slight increase in performance. This effect is known as supercompensation. Supercompensation therefore describes the (rather limited) phase of increased performance as well as the effect as such. The body tries to arm itself a little against the type of demand or overload. The supercompensation phase begins immediately after complete regeneration, the performance level slowly rises by a small amount up to a maximum increase, only to then slowly fall again and finally return to the original performance level. Now, of course, you would want to place the next training session directly at the maximum level of supercompensation in order to utilise this effect again at a slightly higher level. And again and again. However, the increase in performance does not increase proportionally without end, but probably follows more of a logarithmic curve, so that the achievable increase decreases further and further towards the end or disproportionately more effort is required for the same increase. The supercompensation curve must therefore be taken into account for optimal training, although this is difficult to predict or calculate in general terms. They are very individual and depend on the type and intensity of training.
The three phases according to the supercompensation theory are thus:
- Training with loss of performance during training,
- Regeneration with recovery to the original performance level
- Adaptation phase with supercompensation, which falls back to zero towards the end of the adaptation phase. There is an excess of anabolic recovery processes here. Supercompensation is reversible within a rather short time.
The mechanisms that lead to the phenomenon of supercompensation have not yet been sufficiently clarified, nor has the mechanism of hypertrophy been adequately explained. One component of this will be the increased storage of glycogen, another an increase in protein synthesis. The increase in protein synthesis within the muscle cells is caused by:
- Anabolic steroids (endogenous or exogenous)
- Microtraumas in the musculature
- metabolic stimuli such as oxygen deficiency
This causes the muscle fibres to hypertrophy: the cross-section increases. Timely replenishment of the glycogen stores in the muscle cells through carbohydrate intake shortens the regeneration time. This resynthesises degraded ATP. A protein-rich diet promotes protein synthesis and leads to a positive net protein balance.
Combination exercises
Bench presses, but above all pull-ups, squats and cross lifts are classic combination exercises in which several muscle groups work simultaneously. This generates a higher output of growth hormones than isolated exercises. At least 40-50% of exercises should be combination exercises.
Endurance sports versus weight training and muscle building
If you are aiming to build muscle and strength, you should not do too much endurance sport, as this does not contribute much to this, but reduces many resources. Running in the days between a three-part split does not optimise muscle building, but delays it. If you value endurance sport, you should limit it to 3 * 20 minutes / week. If there is steady progress in strength despite endurance training, it can be assumed that endurance sport is not interfering (too much).
Further information
Incomplete short range movements, if performed in rather short sarcomere lengths, lead to a loss of training effect. They reduce the TUT at the same speed and tend to shorten the muscles even more.
Swing has no place in strength training. Resting for more than two to three minutes between sets is counterproductive. Holistic exercises such as squats are superior to machine exercises. Once the muscles are sufficiently warmed up, the contraction speed can be increased: explosive movements activate more fast-contracting muscle fibres. However, the increased risk of injury must also be borne in mind. The associated eccentric contraction can be significantly slower. Rest in terms of exertion is not recommended during strength training. If strength deficits become apparent, they should be addressed quickly.
Smoking reduces the oxygen supply via the supply of carbon monoxide and therefore performance and training success. Alcohol reduces testosterone and even leads to a decrease in muscle mass.
After training, carbohydrates and proteins should be consumed. If the carbohydrates are not consumed, the body breaks down proteins in order to produce glucose from protein via gluconeogenesis. A recommended ratio between carbohydrates and proteins after training is 4:1. However, according to a meta-study by Schoenfeld et al. (2013), it is not the timing but the daily amount of protein that is decisive for muscle growth. Regular and sufficient sleep at regular times is supportive, breaks in this regularity are detrimental. The last sport and the last caffeine should be approx. 4 to 6 hours before sleep. Consuming too much sugar leads to blood sugar spikes that make you feel full and prevent or reduce the absorption of valuable nutrients.
Sufficient drinking supports protein metabolism, especially important when large amounts of protein are consumed. Roughly speaking, 5-6 times 25-30 g of protein per day are recommended, or according to other recommendations 4 times a day 0.4 g per kg of body weight per meal. Formulas are usually used that take body weight into account, with most figures being 1.4 – 2 g of protein per kg of body weight, which is around twice as much as the 0.8 g per kg of body weight recommended by the German Nutrition Society for people who do not do muscle-building strength training, but at best moderate recreational sport. A slightly higher protein intake of 1 g is recommended for people aged 65 and over in order to better counteract sarcopenia. A protein intake in excess of this amount harbours risks, for example for the kidneys.
Intensity levels of the training
The aerobic threshold (also known as lactate equivalent or baseline lactate) is defined as the intensity of exertion above which the body can no longer produce energy purely aerobically during prolonged exercise. For most people, it is 2 mmol/l of lactate, while 1-2 mmol/l are produced at rest.
In spirometry, the aerobic threshold corresponds approximately to the VAT (Ventilatory Anaerobic Transition), the threshold at which the metabolism becomes partially anaerobic. Up to this point, fats and KH are used to generate energy. The IAS (individual aerobic threshold) as the point of the first lactate increase can be slightly different. After this point, only carbohydrates are used for further energy production. Depending on the training status, more or less oxygen is supplied. If the oxygen supply is good, a lot of energy is generated and only water and CO2 are produced as metabolic products, which can be easily removed as blood gases. If the oxygen supply is poor, little energy is generated and a lot of lactate is produced, which increasingly acidifies the muscle; the lactate can be metabolised in real time by the brain and liver up to a certain level (steady state). The steady state depends on how good the oxygen supply and the penetration of the muscles with the smallest vessels is (capillarisation). Both parameters are improved by regular endurance training (at least 3 / week), as is the lactate metabolism performance of the liver.
With further or longer-lasting performance demands, the system decompensates, the anaerobic threshold is exceeded, which is the highest possible exercise intensity that can be maintained without increasing acidosis; it is 4 mmol/l of lactate in the blood, but may differ slightly from this as IANS [Individual Anaerobic Threshold]. The over-acidification of the muscle irritates nerve cells, the muscle begins to ache, fatigue and cramps can occur. The anaerobic threshold can therefore not be exceeded for long, especially as it also has an influence on motivation. This phase can therefore only cover a final spurt, for example. The recovery phase required afterwards is relatively long.
Some of the adaptive benefits that trained athletes exhibit:
Higher VO2max, improved capillarisation of the muscles, higher possible lactate concentrations, shortened regeneration phase, i.e. earlier recovery of performance capability, increased perspiration to avoid performance-inhibiting heat build-up due to metabolic heat (exothermic metabolism) in the muscles.
Through endurance training, these parameters can be shifted in the direction of competitive athletes, with the optimum being a continuous stimulus in the area of the steady state (anaerobic threshold), where rapid and sustainable adaptations and performance gains can be achieved. While a competitive athlete can still generate aerobic energy at 80% VO2max, the untrained athlete can only do so at 50% VO2max. This measured variable is usually related to body weight as ml/min/kg and can be determined in clinical tests as the oxygen uptake at the time the performance requirement is cancelled. The standard value is gender-dependent and decreases with age.
Other measured variables are:
– carbon dioxide release
– respiratory minute volume VE as the product of tidal volume and respiratory rate. As with the regulation of blood flow, the depth of breathing, i.e. the tidal volume, and then the respiratory rate are increased with increasing performance requirements. In the case of obstructive or restrictive respiratory diseases, these patterns may be altered.
– the respiratory quotient RER (respiratory exchange rate) as a quotient of VCO2 and VO2, which is between 0.7 for pure fat burning and 1 for pure carbohydrate burning. At high exercise intensities, the quotient exceeds 1
– the anaerobic threshold AT
determined by the lactate test – the ventilatory threshold VT or VAT (Ventilatory [anaerobic] Threshold), i.e. the point from which respiration increases non-linearly and metabolic acidosis must occur, which occurs from a further kink point RCP (Respiratory Compensation Point) in the RER because H+ must be increasingly exhaled.
In order to test performance according to these parameters, the following conditions must be met:
– last infection at least 14 d ago, good general state of health and motivation
– 48 h alcohol abstinence
– at least 48 h after last exhaustive exercise
– caffeine and nicotine consumption as usual
– temperature 18°-24°, humidity 30%-60%.
training zones
The traditional training zones are divided into 10 per cent increments as follows:
- Health zone (regeneration, compensation, very easy): 50-60% of HFmax, strengthens the cardiovascular system, good for beginners
- Fat-burning zone (basic endurance training 1, easy): 60-70% of HFmax, strengthens the cardiovascular system, improves fitness, best fat-burning effect
- Aerobic zone (basic endurance training 1-2, moderate): 70-80% of HFmax, improvement of breathing and circulation, improvement of aerobic fitness, best promotion of endurance
- Anaerobic zone (basic endurance training 2.intensive): 80-90% of HFmax, accumulation of oxygen debt, improvement of lactate tolerance, short-term training intervals for competitive athletes, training for maximum performance gains. The beginning of the anaerobic threshold is the „lactate threshold„
- Red zone (competition zone, competition-specific endurance training, maximum): from 90% of HFmax, improvement in maximum performance and speed, dangerous for recreational athletes
Anaerobic training only burns KH, instead of the red muscle fibres, the white muscle fibres are mainly recruited. Sprints, competitive sports and strength training are typical, while light endurance sports such as jogging and swimming are typical of aerobic training. The goals of aerobic training are to build muscle and increase performance instead of burning fat and increasing endurance.
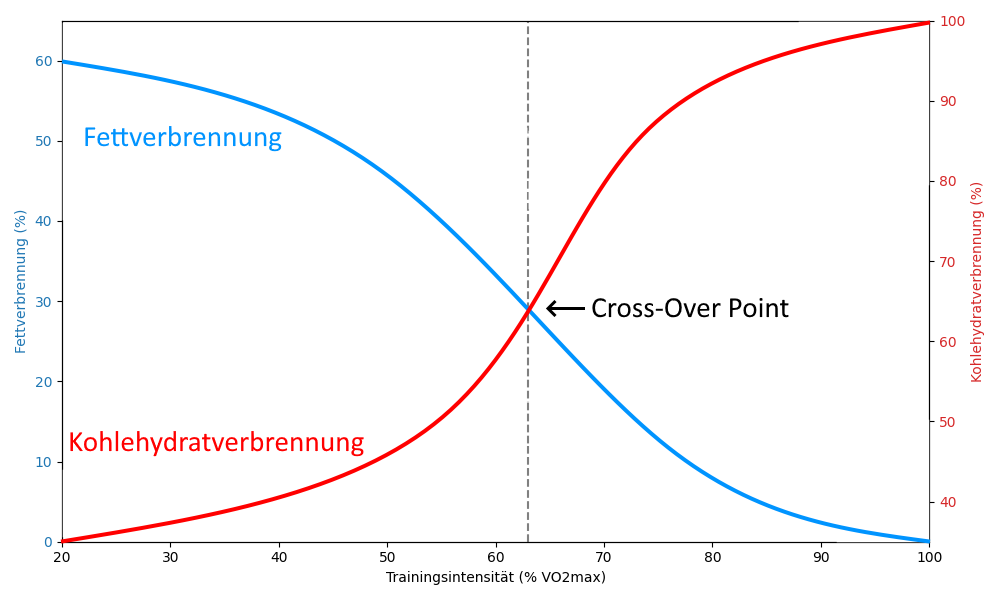
Fat and carbohydrates
The body is fundamentally capable of burning fat or carbohydrates, or both in varying proportions, to generate energy. The respective proportion depends mainly on the intensity of the training, but also on the duration of the training. Trained individuals also burn fat more and earlier than untrained individuals, which makes sense because the body’s fat reserves are much larger than its glucose reserves (or glycogen as a storage form). If the ratios are plotted according to training intensity, the following curve is obtained for endurance sports. The COP (cross-over point) is the intersection of the two curves for fat consumption and carbohydrate consumption.
Applied according to the duration of training, the following picture emerges, whereby RCOP denotes the intersection of the two curves for fat consumption and carbohydrate consumption:
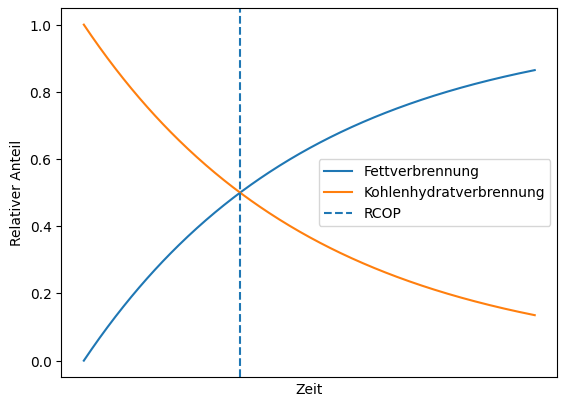
Studies show that at low training intensity (50–60% of VO2max), it takes 3 to 5 hours to reach RCOP, not least because carbohydrate stores are depleted more slowly at this intensity. At over 70% VO2max, it takes 2–3 hours to reach RCOP, although at 80% and above, carbohydrates are predominantly burned even during longer training sessions. RCOP then either does not occur at all or occurs with a significant delay. The athletes‘ level of training also plays an important role, especially in endurance sports, resulting in the following schematic picture:
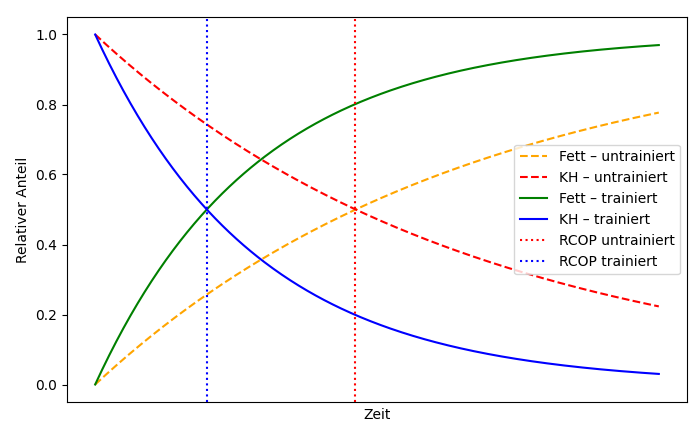
In strength training, the RCOP is usually not reached for several reasons. On the one hand, the training sessions are usually not long enough. Even after 90 minutes of strength training, the carbohydrate stores are unlikely to be depleted. However, if strength training is carried out over a longer period of time, i.e. over many months, the increased muscle mass raises the body’s basal metabolic rate at rest, which also contributes to increased fat burning. After all, one gram of muscle mass requires 6–10 calories per day to maintain itself, even without physical activity, i.e. at rest. In contrast, fat tissue only needs 2 calories per gram. Carbohydrate stores are mainly located in the muscles (300–500 g glycogen) and liver (80–150 g glycogen), totalling approximately 400–650 g glycogen (the stored form of glucose, a much more complex molecule). in untrained individuals, this is more likely to be 300–400 g in the muscles, while in trained individuals it is more likely to be 500–600 g in the muscles, sometimes even more. 1 g of glycogen (or glucose) corresponds to approximately 4 kcal, which means that glucose stores amount to approximately 1600–2600 kcal. Glycogen is the intracellular storage form, whereas glucose is mainly found in the serum, but only in amounts of around 4 g. Glycogen is therefore more readily available than glucose because it is already present in the body.
Carbohydrate loading, practised in advance by endurance athletes for competitions lasting 90 minutes or more, provides an additional 10–15% storage capacity, i.e. up to approximately 700 g. Trained individuals also replenish their stores more quickly than untrained individuals. With the equivalent of 1 kcal ≈ 1.162 Wh, this results in a storage capacity of 1860–3020 Wh of work equivalent or energy. To reach values of 1000 kcal, i.e. around half of the glucose storage, a strength athlete would have to perform whole-body training for around 2 hours at a time (without set breaks) to achieve values of 1000 kcal, i.e. around half of the glucose reserves. This requires 600 to 1000 kcal, which corresponds to around 700 to 1150 kcal or 175 to 290 g of glucose. Of course, these values vary depending on body mass and gender.
Anaerobic metabolism produces two ATP molecules from one glucose molecule, whereas aerobic metabolism (respiratory chain and citric acid cycle) produces 30–32 ATP molecules. Glycogen produces 3 ATP molecules anaerobically or 33–34 ATP molecules aerobically. Fat is more energy-rich (9 kcal/g) than glucose/glycogen and is mainly stored in subcutaneous fat tissue and visceral fat tissue. During physical activity or fasting, fatty acids are released from the fat tissue into the blood, from which the cells can produce ATP.
The energy stored in fatty tissue is significantly higher than that stored in glucose reserves: an athlete weighing 75 kg with a BMI of 21 and 12% body fat, corresponding to approximately 9 kg of fat, stores around 63,000–67,500 kcal, which is approximately 73,200 to 78,435 Wh or around 75 KWh. which is roughly the energy required to power a small electric car at maximum performance for one hour, or to drive a small electric car at 130 km/h on the motorway for three hours.
Karvonen and the need to consider the resting heart rate
In Karvonen’s simplified model, only three zones are defined, but they are dependent on the resting heart rate. They are the proportion of the heart rate reserve by which the heart rate is increased during training:
- Untrained: 0.5
- extensives, lockeres Ausdauertraining: 0,6
- Intensive endurance training: 0.8
The need to include the resting heart rate is illustrated very well by the following example: of two men who are both 70 years old, 185 cm tall and weigh 70 kg, let
- Person A has been an ambitious cross-country skier for 50 years, which is why his resting heart rate is 40.
- Person B has always been completely unathletic and physically inactive throughout; he may also suffer from some cardiovascular/pulmonary health problems, which is why his resting heart rate is 100 bpm
According to the traditional model, B is in the fat-burning zone T0.6 even when sleeping at night, while A has to do an activity that increases his pulse by a full 60 beats for a pulse of 100 bpm, one and a half times the resting pulse, which fits T0.6 much better.
A will not even reach heart rate 100 during heavy but interrupted activities such as strength training, while B would remain in training zones T0.6 or even T0.7 for a long time during the same activities due to a poor recovery heart rate. Apart from this, the exercise-induced increase in heart rate by 60 beats compared to the resting heart rate, which first brought A to 100 bpm, would perhaps no longer be possible for B (Hossak), and on the way there he would certainly experience angina pectoris heart attack-like symptoms, which would force him to stop exercising immediately.
Here you can calculate the HRmax with details of the training zones according to Karvonen that depend on the HRmax and resting heart rate.
Muskelkater (Munich Consensus Statement 1b)
Muscle soreness (actually -catarrh) is the pain phenomenon that occurs as a result of mechanical damage to Z-discs. High loads, especially eccentric contraction (this can be significantly stronger than concentric contraction), particularly braking movements, cause tears in the Z-discs. This can also be caused by intensive stretching. This is followed by an aseptic inflammatory process with migration of water and formation of a slight oedema. Proteins continue to degenerate and autophagy sets in. As the muscle fibres have no pain receptors (but do have stretch receptors), pain only occurs with a delay of 12 to 24 hours when certain substances involved in the inflammatory process escape from the sarcomere and come into contact with nerve endings.
The explanation that it is a lactic acid deposition is outdated: the half-life of lactate is 20 minutes, so the delayed onset of muscle soreness does not match its breakdown. In addition, muscle soreness mainly occurs during strength training or intensive stretching, during which little lactate is produced. However, the 400m run, which produces a lot of lactate, generally results in less muscle soreness.
Stretching before or after intensive muscle strain has no great influence on the development of muscle soreness. However, a good warm-up reduces the risk of injury and improves performance.
Gentle (not vigorous) massages reduce the expected muscle soreness a little, as they promote blood circulation; vigorous massages are counterproductive, as they mechanically irritate the muscles too much. Heat treatments relieve the pain and promote healing because they stimulate blood circulation. Preventive or post-exercise large doses of protein alleviate muscle soreness, with BCAA (branched-chain amino acids) having the best effect.
If very long sarcomere lengths are trained so intensively that muscle soreness occurs, longitudinal muscle adaptation can occur, i.e. the muscle increases the number of serial sarcomeres (this is usually a 4- or 5-digit number) in order to be better able to cope with a further, similar load, as the larger number of serial sarcomeres makes the working range (in terms of sarcomere length) more favourable, i.e. it shifts from the upper limit slightly below 5 µm towards the medium length (about 2.5 µm), which has the advantage that the force that can be deployed becomes greater in terms of the force-length function. An example of this for less mobile people is the right-angled uttanasana or the deep cross lift, which affects the ischiocrural group, as it has to hold a large partial body weight (in the case of cross lifting plus an external weight) at a very large sarcomere length.
Muscle strength
The physical performance that a muscle delivers. The maximum muscle performance (efficiency) is usually of interest, which is why this is often referred to when we talk about muscle performance. Since power in the physical sense is the quotient of work (force * distance) and time, halving the time in which a weight is lifted, for example, leads to twice the required power. This is therefore a hyperbolic dependency with a singularity. On the other hand, a greater weight can be lifted with greater time input. This process finds its natural limit at the maximum tendon force that a muscle can generate and its conversion according to the lever ratios as well as the relation of the movement to the gravity direction according to the sine function. The extreme values of this dependency lie on the one hand, for example, in arbitrarily fast bending of the elbow joint without external weight (utilisation of maximum power in terms of speed), so that the muscles only work against the inertia and gravity of the arm itself (other factors such as air resistance or internal tension of tissues incl. antagonistic musculature should be neglected here) and, on the other hand, in gravity movements that are performed firstly concentrically (F1) at minimum (fixed) speed, secondly statically held isometrically (F2) or thirdly eccentrically (F3) at a defined speed, whereby the forces that can be exerted can be estimated against each other so that F1 is smaller than F2 and this in turn is smaller than F3. The isometric case does not fit into the classical understanding of work, as no mass is moved in any way. Nevertheless, the muscle with its constantly alternating fibre bundles „always applies the force to hold the weight. In addition to these considerations, it must be noted that the maximum force that a muscle can develop depends on the current sarcomere length. Therefore, strictly speaking, due to the maximum force depending on the current sarcomere length (see the force-length function), when measuring the power, it must be determined at which sarcomere length or in which interval of sarcomere lengths (or joint angles) the sarcomere length is to be measured. joint angles) the power is measured. A third type of performance requirement can be identified as the maximum number of repetitions that a muscle can perform with significantly submaximal effort, as is the case with running, whereby the influence of factors external to the muscle such as respiration, oxygen transport, quality of the air we breathe (think of higher altitudes), storage capacity for various substances and the current level of the stores become significantly more influential. The more a performance requirement shifts towards „endurance performance“, the more the factors external to the muscles become important. In extreme cases such as multi-day races, for example the Self-Transcendence 3100 Mile Race, the body’s metabolic performance, and mainly the intestinal area, plays a greater role than the classic muscle performance, as this race consumes around 10,000 kcal per day over more than 40 days, so that the highest possible number of calories must be absorbed and utilised every day.
In practice, the performance of a muscle is not constant over a training session, but decreases noticeably and measurably at the end of a training session if it only has a certain minimum intensity. On the other hand, the performance achieved over a certain period of time is directly proportional to the energy used. So if you try to reach a speed of 100 km/h as quickly as possible with a car in a model example, you can be sure that you have used as much energy as possible, i.e. consumed as much fuel as possible. In this example, further resistance must be continuously overcome in order to drive, mainly rolling and air resistance. Extremely slow acceleration does not result in optimum fuel consumption (minimum energy input), as the integral over the other resistances exceeds any energy input for acceleration over time. There is therefore a certain constant but moderate acceleration behaviour which is energy-optimal. For the weight to be lifted above, this means that the weight that can (just barely) be held is greater than any weight that can still be actively lifted, but an arbitrarily slow lift also does not lead to optimal behaviour, but rather fatigues the muscle by exhausting the local energy reserves of the muscle to such an extent that muscle failure occurs.
Muscle performance
The maximum physical power (muscle power) that a muscle can produce. This value clearly depends on the time over which the performance is to be achieved.
Muscle failure
There are three forms and phases of muscle failure, which typically occur one after the other with continued intensive exercise and can therefore be assessed against each other.
- Concentric muscle failure: the inability to continue concentric contraction
- Isometric muscle failure: the inability to continue holding an isometric contraction
- Eccentric muscle failure: the inability to continue to contract the muscle eccentrically in a controlled manner, for example to slowly release a weight to the ground with a delay.
In heavy weight training, all three stages of failure would normally be provocable one after the other if in the last repetition, in which concentric contraction is no longer possible after a certain point (concentric failure), the isometric contraction is held for as long as possible and, after isometric failure has occurred, an attempt is made to perform the eccentric contraction as slowly as desired, which will lead to eccentric failure.
The three stages of muscle failure can be easily understood with the following simple experiment, which provokes the stages of muscle failure one after the other: lift a weight in the order of 80-90% of the maximum strength, for example in the sense of a bicep curl, again and again for as long as possible. At some point, the energy reserves stored in the muscle will be too low to lift the weight again. The fuel supplied via the bloodstream cannot cover the energy requirement needed for this weight in real time anyway, so the barbell visibly stops „halfway“. The phenomenon observed is concentric muscle failure. If you now try to hold the weight in this position, you will notice that this is also no longer possible at some point, then isometric muscle failure occurs. The attempt to lower the weight as slowly as desired will also come to an unwanted end due to increasing exhaustion, eventually leading to eccentric muscle failure: the inability to lower the weight with eccentric muscle contraction in a controlled manner. The reason for this behaviour naturally lies in muscle physiology: even holding an object against gravity requires constant muscle work by alternating fibre bundles and thus consumes energy. In contrast to what can be realised in technical systems, there is therefore no mechanism by which the muscle „locks in“, so to speak, so that holding it no longer requires any further energy. In the sense that muscle failure represents a supply that is no longer possible in real time, it has a kind of analogy with the aerobic threshold, at which oxygen can no longer be absorbed according to consumption. The above experiment also shows that a concentrically achievable force is smaller than the isometrically achievable eccentric force, whereby the greater the concentric or eccentric contraction speed, the greater the difference between the achievable force and the isometric force.
Some training instructors claim that more or less frequent muscle failure is necessary for effective training. However, the only certainty is that „suprathreshold stimuli“ are necessary. A certain amount of caution is required when it comes to training to muscle failure: the muscle must be very well warmed up for this and the risk of injury is significantly increased anyway in the mode in which the weight can no longer be controlled well. In addition, training to the point of muscle failure places great demands on the CNS. For locally exhaustive training, 48 hours of recovery is the lower limit.
Eccentric muscle failure
the last form of muscle failure, which occurs after the other two forms or stages: the inability to perform an eccentric contraction at a slow speed in a controlled manner. For an explanation and example, see muscle failure.
Isometric muscle failure
the middle of the three forms or stages of muscle failure: the inability to maintain a given isometric contraction. For an explanation and example, see muscle failure.
concentric muscle failure
the first form of muscle failure: the inability to continue concentric contraction, for explanation and example see muscle failure.
longitudinal muscle adaptation
The number of sarcomeres in a myofibril is not a fixed value that remains constant throughout life. The use of the movement apparatus does have an influence on this. For example, muscles that are immobilised for a short sarcomere length over a longer period of time (usually as a result of trauma) reduce the number of serial sarcomeres (i.e. those that lie one behind the other in the myofibril), thus reducing their number. Similarly, more frequent intensive use of the muscle in longer sarcomere lengths is likely to increase the number of serial sarcomeres. This is referred to as longitudinal muscle adaptation in contrast to the long-known growth in muscle thickness through training, in which the number of parallel sarcomeres increases. The consequences of the (reversible, see above) longitudinal muscle adaptation result from the fact that more sarcomeres now perform the same work in series than before and, as far as is known today, are consistently positive:
- Favourable shift of the working range of the individual sarcomeres in terms of the force-length function
- Slight increase in muscle performance
- Slight increase in the maximum force
- Slight increase in maximum contraction speed
- Slightly reduced susceptibility to injury
For examples of postures that typically cause longitudinal muscle adaptation, see the explanation under muscle soreness.
It is now known that titin triggers longitudinal muscle growth via the mTORC1 signalling pathway when subjected to significant stretching.
Pre-activation
Pre-activation refers to the training principle in which the relevant muscles are challenged with a minimum intensity (medium or higher) shortly before a planned performance (around 5 minutes) so that they are more efficient for the planned activity. Pre-activation can be performed quickly or isometrically. In the literature, you can find information on intensity between 30% and 70% of maximum strength.
Several mechanisms are responsible for the effect:
– Increased phosphorylation of the myosin chains enables better and faster contractions
– Increased motoneural excitability
– A more favourable muscle flexion angle increases the possible force in feathered muscles Hopping, skipping, jumping rope, pulling exercises with fitness bands, isometric maximum contractions in the 3*10-15 s or one minute total range are favourable.
The time of optimal pre-activation should be somewhere between 30 seconds and a few minutes. If the pre-activation is too intense, the muscle is already a little fatigued when the actual performance is to be performed. If it is too far away from the planned performance, the pre-activation effect has already partially subsided.
Sarcopenia
The effect that the muscle mass of a person decreases with age with otherwise reasonably constant parameters is known as sarcopenia. This effect is in the order of 1-2% / a. Sarcopenia necessarily entails restrictions in quality and development of life, as well as increased risks, such as falls or bone fractures, which occur much more easily in the presence of osteoporosis favoured by sarcopenia. Depending on the extent, everyday activities such as lifting and moving objects, various activities requiring strength and even locomotion, especially faster walking or climbing stairs, can also be significantly impaired by sarcopenia. In one study, the risk of death was found to correlate with the degree of sarcopenia. According to current knowledge, the best method against sarcopenia is strength training of any kind. It is of no great importance whether the exercises are performed against external resistance such as weights or resistance bands or, as far as possible, against the body’s own weight. However, this should be introduced after a long period of abstinence from sport.
Dynapenia
Dynapenia refers to the loss of muscle strength, not due to underlying neurological, metabolic or muscular pathology, but as part of the physiological processes and lack of maintenance associated with ageing. It has been observed that muscle performance declines first and most noticeably, followed by muscle strength (dynapenia) and then muscle mass (sarcopenia). According to one experiment, the effect of sarcopenia is only responsible for 10% of the decline in strength. Muscle strength also depends to a large extent on the control of the muscle by the CNS and on the muscle’s internal ability to supply energy. Statistically speaking, the central activation of the muscles continuously decreases in non-exercisers, so that in a corresponding study with very elderly people (85–97 years), central activation was found to decrease by 69–93% (average 81%). This means that the loss of strength was five times greater than the loss of muscle mass (sarcopenia). Various neurological phenomena contribute to dynapenia:
- Disorders affecting the integrity of the white matter in the brain
- increased intracortical inhibition
- Decrease in the size of neuron bodies
- cortical atrophy
- Reduction in the length of myelinated fibres
- Changes in various neurotransmitter systems
- lower expression of neurotrophic factors
- in the spinal cord: decrease in the ability of the motor units to discharge, reduced number and size
- in the muscle: myosteatosis
- in the muscle: decreased expression of sarcoplasmic reticulum proteins, resulting in adverse electrolyte changes
- in the muscle: adverse changes in myosin
- in adipose tissue: cytokine production affecting the muscles
This leads to a decrease in both cortical excitability and neuroplasticity. The neurological changes in the CNS are therefore an extremely important factor in dynapenia. Mental imagery (imagining strong muscle contractions) can replicate the effects of real training to a certain extent. One study showed a loss of functioning motor units in older people. However, it is also known that decades of aerobic exercise provide good protection against the loss of motor units that is otherwise commonly associated with ageing.
Predisposing factors for dynapenia are:
- declining physical activity
- Existing weakness; sluggishness, which is often an expression of limited capabilities; unwanted weight loss; low cardiopulmonary endurance and local muscle endurance; insufficient protein intake
- Age over 80 years
To date, there is no widely recognised test or score for dynapenia. However, it is considered certain that resistance training/strength training is an excellent way to counteract dynapenia.
Myogelosis
This circumscribed hardening of the musculature is considered to be the result of overuse or incorrect strain as well as stress. It is a fixation that appears histologically degenerated like wax. Palpation of myogelosis is characterised by hardened nodules and painful pressure. The shoulder/neck and back are most commonly affected. Myogelosis is regarded as the palpatory equivalent of trigger points, in which the pressure pain is in the foreground. Some authors equate myogelosis with muscle hard tension, others differentiate in such a way that myogelosis is described in small volumes and muscle hard tension affects the entire muscle. A further difference is that hard muscle tension reacts to stretching and local anaesthetics, whereas myogelosis does not.
Arthrogenic muscle inhibition (AMI)
Arthrogenic muscle inhibition (AMI) is a persistent presynaptic reflex inhibition that originates from injured joints and is understood as a protective reaction of the body against further damage to the joint. It originates from mechanoreceptors of the joint (Ruffini endings, Pacinian capsule, Golgi tendon organ, free nerve endings) and acts via inhibitory interneurons, which lead to reduced contraction. Since 2000, there have been many studies on this, particularly in the area of the knee joint, especially on damage and ruptures of the ACL, arthrosis interventions in the knee joint and other conditions. At 84%, the quadriceps is the predominantly examined muscle, followed by the ischiocrural group at approx. 9%. It is known from the AMI of the quadriceps that various knee injuries and interventions restrict the contraction force. The effect depends on factors such as the extent of joint effusion, the time elapsed since the damage and the angles still achievable in the knee joint. Inflammation, instability of the joint and damage to the afferents also interfere. AMI is considered to be one of the important mechanisms that hinder rehabilitation after joint injury, reducing or limiting the use of muscles important to the joint. Pathomechanically, in addition to reflex-like muscle inhibition, there are also deficits in proprioception as well as neuroplastic compensations in the brain, which together lead to reduced motor performance of the muscles relevant to the joint. It must be assumed that without elimination of the AMI, complete rehabilitation of the joint remains difficult.
Stretching
Stretch: The connective tissue structures of the muscles are only stressed when they are stretched considerably, their undulating fibres are stretched (creep effect) and the extended stretching capacity can be used for up to an hour. The muscle fibres have stretch receptors (muscle spindles) that report the state of stretch to the brain. Stretching the muscles can lower the stimulus threshold of the receptors. Regular stretching as well as strength exercises lead to an increase in the stabilising titin.
A distinction is made between mobility as the amplitude (angular range in the joints) that can be actively achieved, whereas flexibility is the passively achievable amplitude.
It is assumed that a stretched muscle has the same maximum force as a shortened one, but over a larger range, in other words: the force-length curve is more elongated because there is optimal filament overlap over a longer range. It is also assumed that a muscle adapts to frequently performed movements and shifts its maximum strength to where it is used most vigorously: Strength training close to maximum length will shift the strength maximum there, strength training close to maximum contraction will also shift this towards maximum contraction and shorten the muscle. It is also obvious that a more stretched muscle has a lower resting tension.
A recent study on mice suggests that stretching inhibits tumour growth, among other things, via a cytotoxic immune response, by around 50% in the study.
Stretching pain
See the page Nerve strain/Stretching pain.
Painful stretching
See the page Nervous system/Stretching pain.
Slackening points and insufficiencies
Slackening points
The passive relaxation point is the degree of stretching of the muscle at which the titin filaments no longer exert force to contract the muscle, which is the case at around 2.8 µm.
The active relaxation point is the point at which the muscle can no longer exert force for further contraction because the myosin heads accumulate in front of the Z-disc; this is the case at around 1.3 µm.
Insufficiencyen
Active insufficiency
State of complete contraction of a muscle, i.e. complete interlocking of actin and myosin filaments, which does not allow any further active concentric contraction and thus movement caused by this muscle itself in a joint, which this muscle covers, although the covered joint or the covered joints themselves are not yet in the end position (reaching a soft-elastic, firm-elastic or hard-elastic movement limit), i.e.
In single-joint muscles, active insufficiency is always pathological; in double-joint (or polyarticular) muscles, it is more the rule.
Passive insufficiency
State of a muscle not being fully contracted, i.e. not actively insufficient (actin and myosin filaments have not fully engaged), but being prevented from further contraction by a lack of flexibility of its (more precisely: one of its partial) antagonist(s).
Passive insufficiency therefore means that a muscle could continue to shorten and move in a joint, but its antagonist(s) prevents this.
In monoarticular muscles, the occurrence of passive insufficiency is usually pathological; in biarticular and polyarticular muscles, it is physiological.
Serial:
M. Sartorius,
rectus abdominis
Parallel:
Biceps Brachii,
Glutaeus Maximus
Finnation:
Semimembranosus (single)
Plantaris (double)
Deltoideus (multiple)
Torque
The most important joints of the movement system have rotational centres (three-dimensional mobility) or rotational axes (one-dimensional mobility). In many joints, regardless of the force exerted (contraction force), the distance between the executing tendon of a muscle and the relevant centre of rotation changes. This means a change in the physical lever arm on which the force is exerted, i.e. the force arm, which changes the torque. The force-length function shown above describes the change in the achievable force over the (Sarmoker) length. However, as movements usually involve rotational movements in the joint, the force arm that changes over the angles also plays a role as a multiplicative factor in the achievable torque.
Myokine
Myokines are special interleukins and thus cytokines (peptide hormones) discovered by Pederson in 2007, which are released in mammals during intensive muscle activity. They are hormone-like (paracrine or endocrine) messenger substances that intervene in various areas, including fat and sugar metabolism, the immune system, inflammation and vascular tension. Around 600 myokines are currently (2025) known, but most of them have not yet been sufficiently researched. Strength training or other intensely strenuous activities in particular release myokines. The effects of myokines known to date are consistently positive and also have a positive influence on various chronic diseases such as arteriosclerosis, diabetes mellitus, neurodegenerative, dementia-related diseases such as Alzheimer’s and other forms of dementia as well as cancer. Interesting details are already available in individual cases.
IL-6
The myokine IL-6 increases almost exponentially with the TUT during intensive concentric contractions. During endurance training, IL-6 increases by a factor of 100 in a very short time. On the other hand, a permanently elevated IL-6 serum value independent of activity indicates pathological events in the body, whereby it has a pro-inflammatory effect, among other things. In RA, for example, there are chronically elevated levels of IL-6, which favours the production of autoantibodies and promotes local inflammation and systemic effects. Antibodies can be used to successfully intervene against IL-6 in RA. In contrast, the effect of such antibodies in PA is detrimental, as IL-6 inhibits the formation of IL-17 and IL-23, resulting in more of these pro-inflammatory interleukins.
IL-15
IL-15, a myokine that was also discovered very early on, also has a pro-inflammatory effect and activates T and killer cells, but is released in large quantities during strength training and related intensive activities and favours muscle growth, helps to reduce body fat and reduces fat storage in the liver. Its effect is positive in diabetes mellitus and cardiovascular diseases and in cachexia. In contrast to IL-6, its release is not stimulated by endurance training.
Irisin
Another myokine, irisin, helps to convert white body fat into brown body fat and is thought to have a positive effect on various forms of metabolic syndrome. Irisin is released not only through intensive physical activity and especially endurance training, but also through muscle tremors in cold. It has a favourable effect on fat and sugar metabolism and reduces bone loss. It also has a positive effect on the synaptic plasticity of nerve cells damaged by Alzheimer’s disease.
BDNF
BDNF is a moykin that can also change the structure of the muscles: it helps to convert slow-twitch fibres (endurance performance) into fast-twitch fibres (strength and speed) and to adapt the innervation accordingly. The result is that intensive and frequent strength training can actually reduce endurance capacity. An unexpected result is that a low level of BDNF (Brain-Derived Neurotrophic Factor) reduces sarcopenia. BDNF is blood-brain barrier-crossing and stimulates a growth factor in the hippocampus that reduces the death of neurons and contributes to the formation of new synapses and
neurons and improves memory and learning ability. BDNF therefore helps prevent dementia and should also be used in cases of depression, anxiety disorders or ADHD. Physical activity is also said to help with depression, anxiety disorders or ADHD with a low serum level of BDNF. In the muscles themselves, BDNF causes a conversion from slow-twitch (type 1) fibres to fast-twitch (type 2) fibres, so that muscles with stamina become muscles with strength. In addition, the absence of BDNF reduces sarcopenia.
Myonectin
Myonectin (C1q-TNF-related protein 15) is reduced in obesity. It is released during exercise. It promotes the cellular uptake of fatty acids and the oxidation of fatty acids in adipose tissue and the liver. There is evidence that myonectin plays a role in diabetes mellitus and obesity.